Antiangiogenic Therapy for Cancer: Current and Emerging Concepts
Angiogenesis is an essential step in the growth and spread of solidtumors-the cause of more than 85% of cancer mortality. Inhibitingangiogenesis would therefore seem to be a reasonable approach to preventor treat cancer. However, tumor angiogenesis differs from normalangiogenesis in that the resulting vessels are tortuous, irregularlyshaped, and hyperpermeable. These abnormalities result in irregularblood flow and high interstitial fluid pressure within the tumor, whichcan impair the delivery of oxygen (a known radiation sensitizer) anddrugs to the tumor. Emerging evidence suggests that antiangiogenictherapy can prune some tumor vessels and normalize the structure andfunction of the rest, thereby improving drug delivery and normalizingthe tumor microenvironment. This normalization effect may underliethe therapeutic benefit of combined antiangiogenic and cytotoxic therapies.This paper reviews current and emerging concepts of the mechanismof action of antiangiogenic therapies and discusses the implicationsof these mechanisms for their optimal clinical use.
Angiogenesis is an essential step in the growth and spread of solid tumors-the cause of more than 85% of cancer mortality. Inhibiting angiogenesis would therefore seem to be a reasonable approach to prevent or treat cancer. However, tumor angiogenesis differs from normal angiogenesis in that the resulting vessels are tortuous, irregularly shaped, and hyperpermeable. These abnormalities result in irregular blood flow and high interstitial fluid pressure within the tumor, which can impair the delivery of oxygen (a known radiation sensitizer) and drugs to the tumor. Emerging evidence suggests that antiangiogenic therapy can prune some tumor vessels and normalize the structure and function of the rest, thereby improving drug delivery and normalizing the tumor microenvironment. This normalization effect may underlie the therapeutic benefit of combined antiangiogenic and cytotoxic therapies. This paper reviews current and emerging concepts of the mechanism of action of antiangiogenic therapies and discusses the implications of these mechanisms for their optimal clinical use.
We have known for nearly a century that tumors induce growth of new blood vessels.[ 1,2] In 1971, Folkman proposed that blocking angiogenesis should be able to arrest tumor growth.[3] A few years later, Gullino demonstrated that cells in precancerous tissue acquire angiogenic capacity on their way to becoming cancerous, and thus proposed that inhibition of angiogenesis be used to prevent cancer.[4] In the past 3 decades our understanding of angiogenesis in general, and tumor angiogenesis in particular, has grown exponentially, culminating in the approval of the first antiangiogenic agent, bevacizumab (Avastin), for the treatment of advanced colorectal cancer and spawning more than 60 clinical trials using a variety of antiangiogenic agents.[5,6] The widely accepted mechanism of action of antiangiogenic therapy is that these agents prevent the growth and metastasis of tumors by inhibiting the formation of new vessels. While this notion holds true, data from randomized clinical trials show that when used as monotherapy, currently available antiangiogenic agents produce modest objective responses and do not yield long-term survival benefit in patients with solid tumors.[7-9] However, when given in combination with chemotherapy, bevacizumab, an antibody directed against vascular endothelial growth factor (VEGF), produced unprecedented increases in survival (5 months) in patients with metastatic colorectal cancer.[10] These data support the earlier predictions of Teicher, who postulated that combined administration of antiangiogenic therapy and cytotoxic (chemotherapy and radiation) therapy would yield maximum benefit because such combinations would destroy both compartments in tumors: the cancer cells and the endothelial cells.[11] However, this theory seems paradoxical. If antiangiogenic agents destroy the tumor vasculature, the delivery of chemotherapeutic agents and oxygen to the tumor would be compromised, resulting in less cytotoxicity and radiation resistance. The emerging concept of vascular normalization provides researchers with a potential resolution to this paradox: this hypothesis posits that judicious application of antiangiogenic agents can improve the function of tumor vasculature and thus improve the delivery of oxygen and drugs to tumor cells.[12,13] Here I review this emerging counterintuitive concept and the currently well-accepted view that antiangiogenic therapy inhibits neovascularization, as well as the importance of optimizing the dose and sequencing of these therapies.


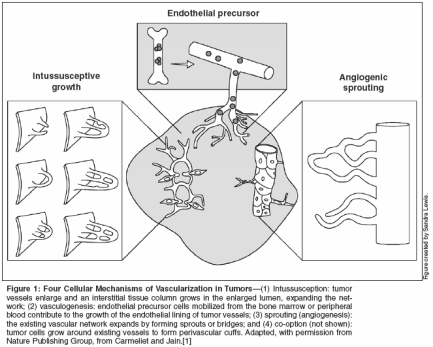
Role of Angiogenesis in Tumor Progression In adults, normally occurring neovascularization is largely limited to female reproductive organs and healing wounds. Otherwise, adult neovascularization occurs as a result of a pathologic situation, such as cancer. Although tumors can initially coopt normal blood vessels to support growth, a new blood supply is required for the tumor to grow beyond 1 to 3 mm. This concept has been firmly established by many supported studies and is well illustrated by a study that found numerous small thyroid gland tumors in individuals who died of other causes.[14] Prior to the development of tumor vasculature, tumors are dormant.[15] For angiogenesis to occur, the effect of proangiogenic factors must outweigh the effect of the antiangiogenic factors. When the balance tips towards proangiogenesis, the tumor undergoes what has been described as the angiogenic switch, which fosters the development of new vessels and allows tumor progression to ensue.[ 16] Further, once angiogenesis is prompted, tumors become invasive locally and systemically. Evidence for the critical and pathologic role that angiogenesis plays in cancer progression comes from studies demonstrating that microvessel density within the tumor-the number of blood vessels in a defined area-is not only prognostic in some tumor types, but also correlates with the development of metastases.[17] Mechanisms of Angiogenesis At least four cellular mechanisms can result in the vascularization of tumors: co-option, intussusception, sprouting (angiogenesis), and vascu- logenesis (Figure 1). Tumor cells can co-opt and grow around existing vessels to form perivascular cuffs. However, these cuffs cannot grow beyond the diffusion limit of critical nutrients, and may actually cause the collapse of the vessels due to the compressive forces generated by tumor cell growth (referred to as solid stress). Alternatively, an existing vessel may enlarge in response to the growth factors released by tumors, and an interstitial tissue column may grow in the enlarged lumen and partition the lumen to form an expanded vascular network. This mode of intussusceptive microvascular growth has been observed during tumor growth, wound healing, and gene therapy.[18-21]






Angiogenesis is perhaps the most widely studied mechanism of vessel formation. During angiogenesis, the existing vessels become leaky in response to growth factors released by normal cells or cancer cells; the basement membrane and the interstitial matrix dissolve; pericytes dissociate from the vessel; endothelial cells migrate and proliferate to form an array or sprout; a lumen is formed in the sprout-a process referred to as canalization; branches and loops are formed by confluence and anastomoses of sprouts to permit blood flow; and finally, these immature vessels are invested in basement membrane and pericytes. During normal physiologic angiogenesis, these vessels differentiate into mature arterioles, capillaries, and venules, whereas in tumors they may remain immature.[12,22-24] Finally, vasculogenesis occurs during embryonic development when a primitive vascular plexus is formed from endothelial precursor cells (also known as angioblasts). Circulating endothelial precursor cells mobilized from the bone marrow or peripheral blood also can contribute to postnatal vasculogenesis in tumors and other tissues.[25,26] However, recent data have questioned the importance and role of these cells in tumor angiogen- esis.[27] One of the current challenges in tumor treatment is to discern the relative contribution of each of the four mechanisms of neoangiogenesis to the formation of tumors to optimize antiangiogenic treatment of cancer.[28]



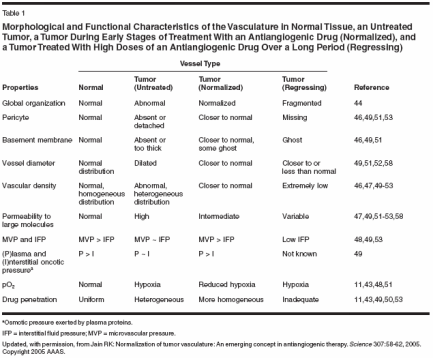
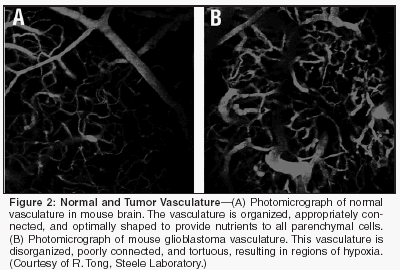
Structure and Function of Tumor Vessels Despite the critical role of blood vessels in tumor growth and metastasis, the structure and function of tumor vasculature is abnormal (Table 1, Figure 2). The organized structure of the vascular network is lost. The system lacks defined arterioles, venules, and capillaries, and connections among vessels are sometimes incomplete. The vessels themselves are irregularly shaped with areas of dilation and constriction. Endothelial cell arrangement is abnormal with the cells separated by wide gaps at one location or stacked on one another nearby. Endothelial cells can lose their reactivity to common endothelial markers.[29] Similarly, the patterns and functions of mural cells are also abnormal. Tumor- associated pericytes demonstrate abnormal protein expression and morphology. Significantly, abnormal pericytes have a loose association with endothelial cells, contributing to the high vascular permeability. The differences between normal and abnormal vasculature are summarized in Table 1. These structural abnormalities result in uneven tumor perfusion and high tumoral interstitial fluid pressure (IFP).[13,30,31] High tumoral IFP is caused in part by tumor vessel hyperpermeability. In normal tissues, the vessel is able to maintain a gradient of fluid pressure from inside the vessel to the outside. In tumors, this gradient disappears and the pressure outside the blood vessels (IFP) tends to become equal to that inside, ie, microvascular pressure (MVP). Similarly, in normal tissues, the colloid osmotic pressure (osmotic pressure exerted by large proteins) inside blood vessels (P) is much higher compared to that outside (I). In tumors, these two become approximately equal due to vessel leakiness. The loss of these pressure gradients between the vessels and the tumor impedes the delivery of large molecular weight therapeutics to the tumor. Uneven tumor perfusion impedes the delivery of all blood-borne molecules, including oxygen and nutrients as well as chemotherapeutics. Tumor vessel hyperpermeability also contributes to sluggish blood flow in tumors, which results in regions of hypoxia and acidosis. Hypoxia contributes to resistance to some drugs and radiotherapy by decreasing the availability of reactive oxygen species. In addition, it can induce genetic instability and upregulate angiogenesis and metastasis genes.[32,33] Furthermore, both hypoxia and acidosis can impede the cytotoxic effects of immune cells infiltrating the tumor. Thus pathologic tumor vasculature results in conditions that protect the tumor from cytotoxic therapy and from host immune cells.


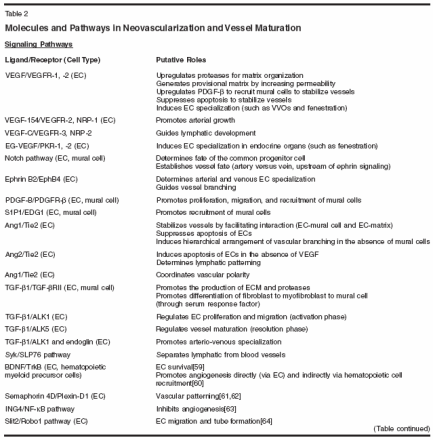
Role of VEGF in Tumor Angiogenesis Numerous pro- and antiangiogenic molecules orchestrate the different steps of vessel formation. These molecules can be grouped into three categories: (1) ligands and receptors, (2) molecules that govern cell to cell interactions, and (3) molecules governing cell to matrix interactions (Table 2). The imbalance of these molecules is thought to promote the pathologic angiogenesis characteristic of tumors. Of the known angiogenic factors, VEGF is the most prominent and critical.[ 1,2,18] VEGF promotes the survival and proliferation of endothelial cells, increases the display of adhesion molecules, and dramatically increases vascular permeability. VEGF expression is elevated in most human tumors and is upregulated by a variety of mechanisms, including hypoxia,[ 33] oncogenes,[5] and hormonal activity.[34] The expression of VEGF correlates with both the extent of angiogenesis and prognosis in many tumor types.[18,35] As a result of its central role in tumor angiogenesis, VEGF has become a prime therapeutic target for antiangiogenic therapy. Mechanisms of Antiangiogenic Therapies The widely accepted mechanism of antiangiogenic therapies is based on the hypothesis that tumor growth is dependent on angiogenesis.[3] Specifically, these therapies are thought to choke the blood supply of tumors, depriving cancer cells of nutrients and subsequently resulting in an indirect cell kill. Indeed, anti-VEGF treatments have proven to be effective in a large number of preclinical studies.[5,6] To date, anti-VEGF treatments-when given as monotherapy-have unfortunately produced only modest responses in patients with solid tumors and have not yet provided significant survival benefit to patients.[9] There are many possible reasons for this failure. For example, as tumors progress, their vessels may become dependent on factors other than VEGF.[36] Thus, blocking VEGF alone may not be enough to induce persistent vessel regression. However, increasing the dose or duration of anti-VEGF treatment might also cause damage to normal tissues (eg, kidney,[ 37] lung,[38] thyroid,[39] and developing bone[40]) and may even result in death due to thromboembolic events in certain subpopulations of patients.[41] As a result, antiangiogenic agents have been combined with cytotoxic therapies (eg, chemotherapy or radiation therapy). Indeed, in a landmark phase III trial, bevacizumab combined with chemotherapy has led to an impressive and significant increase in survival in patients with advanced colorectal carcinomas.[10] At the same time, a phase III trial combining bevacizumab and chemotherapy has not shown a significant survival advantage in breast cancer patients,[ 42] although it did show a significant increase in response rates. These positive and negative survival results with combination antiangiogenic and chemotherapy mimic preclinical findings of augmentation vs antagonism of the efficacy of cytotoxic therapies by antiangiogenic agents.


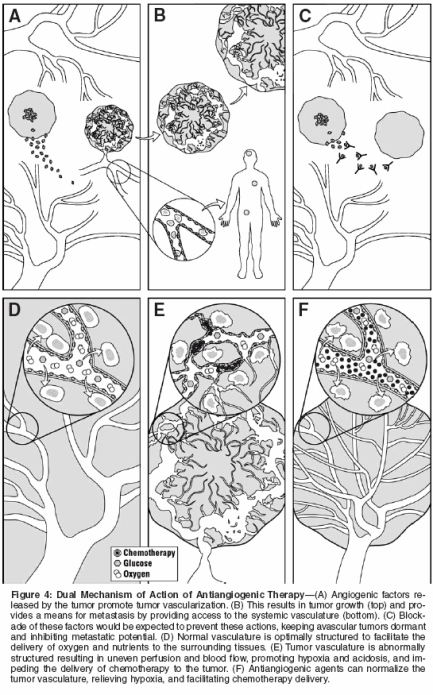
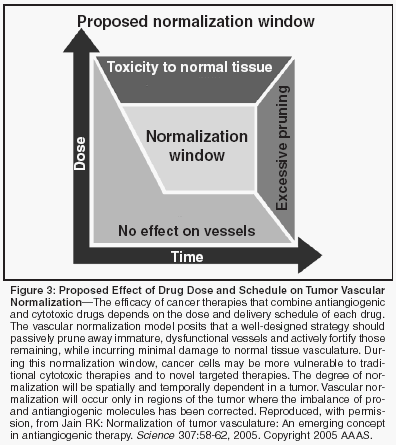
To reconcile these contrasting effects of antiangiogenic therapy, in 2001 I proposed another mechanism of antiangiogenic therapy-normalization of pathologic tumor vasculature by antiangiogenic therapy[12]-and have recently summarized compelling evidence in support of this new concept.[ 13] This theory proposes that normalization occurs because antiangiogenic agents transiently restore the balance of pro- and antiangiogenic factors. By normalizing the structure and function of the tumor vasculature, chemotherapeutic agents and oxygen can be more effectively delivered to the tumor, resulting in improved chemotherapy delivery and radiotherapy sensitivity. While this mechanism is a relatively new concept, it appears to explain the pronounced clinical benefit observed with blockade of VEGF in conjunction with standard chemotherapy.[10] Of interest, there is now increasing evidence in the literature for vascular normalization as a result of VEGF blockade, both in preclinical[11,43- 52] and clinical studies.[53] A clinical study in patients with rectal cancer showed that bevacizumab decreased tumor microvascular density, vascular volume, and IFP.[53] The lack of a concurrent decrease in the uptake of radioactive FDG in these tumors suggested that the normalized vessels were more efficient at delivering these agents to the tumor parenchyma when treated. The reductions in tumor vascular density and permeability were consistent with another study in patients with colorectal cancer treated with an inhibitor of the VEGF receptor tyrosine kinase 2.[54] Unfortunately, in the latter study, there were no measurements of IFP or vessel wall structure to draw any conclusions about vessel normalization. It is important to note that following antiangiogenic drug delivery, a specific time window exists during which vascular normalization occurs. In a murine study that evaluated an antibody to VEGF receptor 2, this normalization window lasted for about 6 days and was characterized by increased tumor oxygenation and vessel pericyte coverage.[51] Consistent with the concept of a normalization window, a recent study showed a transient increase in tumor oxygenation and radiation response of a murine fibrosarcoma when animals were treated with thalidomide (Thalomid), an agent that blocks VEGF and bFGF signaling.[43] Thus, future research must focus on identifying the optimal timing of administration of antiangiogenic agents with chemotherapy or radiation to maximize the benefits that may be attained during this crucial time period (Figure 3). Finally, emerging data show that some cancer cells harbor the receptors for growth factors modulating angiogenesis, and thus may be a direct target of some antiangiogenic agents. For example, some cancer cells express VEGF receptors,[55-57] and ligation of these receptors induces tumor cell proliferation.[57] Blockade of these receptors inhibits VEGFinduced proliferation, providing a mechanism by which some antiangiogenic agents could have direct inhibitory effects on tumor cells. In summary, current data support two key mechanisms of antiangiogenic therapy: (1) inhibition of new vessel formation inside the tumor, and (2) pruning nascent vessels and normalizing the remaining tumor vasculature (Figure 4). Clinical data to date suggest that the latter mechanism may be a key driver of the clinical benefit of combined antiangiogenic and cytotoxic therapies. Future Directions in Antiangiogenic Therapy Angiogenesis is a complex process involving an increasing number of molecular players (Table 2). In addition, there is a likelihood of redundancy in the system and various factors may be critical at different times during vascular development. Because of its critical role in tumor angiogenesis, current therapy has focused largely on blockade of VEGF. Future angiogenic therapy may target VEGF in addition to other angiogenic factors. For example, it has recently been found that the anti-HER2 antibody trastuzumab (Herceptin) blocks multiple angiogenic pathways[58]; combination bevacizumab and trastuzumab therapy is currently in clinical trials. As these combination antiangiogenic therapies are developed, it will be important to balance the benefit of complete angiogenic blockade with any safety concerns that may arise from a potentially too-effective blockade (eg, impaired wound healing, adverse effects in kidneys, thyroid, lung, brain, and heart). Judicious administration of antiangiogenic agents may provide a viable strategy in which multiple pathways are targeted but the blockade does not induce toxic effects. The optimal dosing of antiangiogenic therapy is another key issue for further research. Preclinical data suggest that the concept of using the maximum tolerated dose may not apply in this setting. In some studies the blockade of VEGF activity resulted in adverse effects on normal vessels in multiple organs.[37-40] In addition, some early dose-escalating clinical studies of VEGF blockade suggest that increasing the dose increases toxic effects, which may be directly related to antiangiogenic activity.[7,8] However, large-scale trials must be completed before any conclusions can be drawn. Finally, as clinical development of antiangiogenic agents continues, it may be necessary to reconsider standard metrics of clinical response. Early data suggest that these agents are primarily cytostatic. Thus a greater emphasis on patient survival relative to objective response may be a more appropriate way to evaluate the efficacy of these therapies. In addition, measurement of surrogate markers for functional blockade of angiogenic factors (eg, tumor IFP, blood flow, permeability, vascular density, pericyte coverage, circulating endothelial cells) is critical to optimize the dose and schedule of antiangiogenic agents and cytotoxic therapies. Conclusions Neovascularization is an important physiologic process that enables the growth and metastasis of malignant tumors. This process is tightly regulated, and a number of proangiogenic factors are required for angiogenesis to occur. Both pro- and antiangiogenic factors represent important potential therapeutic targets in cancer therapy. Of these factors, VEGF is the most potent and predominant. Emerging clinical data show that anti- VEGF therapy inhibits tumor growth and metastasis by inhibiting angiogenesis and by normalizing tumor vasculature. This latter effect is an intriguing area of study that may lead to the optimization of combination antiangiogenic and cytotoxic therapy.
Disclosures:
Dr. Jain is a consultant to AstraZeneca.
References:
1. Carmeliet P, Jain RK: Angiogenesis in cancer and other diseases. Nature 407:249-257, 2000.
2. Ferrara N, Hillan KJ, Gerber HP, et al: Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nat Rev Drug Discov 3:391-400, 2004.
3. Folkman J: Tumor angiogenesis: Therapeutic implications. N Engl J Med 285:1182- 1186, 1971.
4. Gullino PM: Angiogenesis and oncogenesis. J Natl Cancer Inst 61:639-643, 1978.
5. Kerbel R, Folkman J: Clinical translation of angiogenesis inhibitors. Nat Rev Cancer 2:727-739, 2002.
6. Hicklin DJ, Ellis LM: Role of the vascular endothelial growth factor pathway in tumor growth and angiogenesis. J Clin Oncol 23:1011-1027, 2005.
7. Cobleigh MA, Langmuir VK, Sledge GW, et al: A phase I/II dose-escalation trial of bevacizumab in previously treated metastatic breast cancer. Semin Oncol 30(5 suppl 16):117- 124, 2003.
8. Yang JC, Haworth L, Sherry RM, et al: A randomized trial of bevacizumab, an anti-vascular endothelial growth factor antibody, for metastatic renal cancer. N Engl J Med 349:427- 434, 2003.
9. Mayer RJ: Two steps forward in the treatment of colorectal cancer. N Engl J Med 350:2406-2408, 2004.
10. Hurwitz H, Fehrenbacher L, Novotny W, et al: Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med 350:2335-2342, 2004.
11. Teicher BA: A systems approach to cancer therapy (antioncogenics + standard cytotoxics → mechanism(s) of interaction). Cancer Metastasis Rev 15:247-272, 1996.
12. Jain RK: Normalizing tumor vasculature with antiangiogenic therapy: A new paradigm for combination therapy. Nat Med 7:987-989, 2001.
13. Jain RK: Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy. Science 307:58-62, 2005.
14. Harach HR, Franssila KO, Wasenius VM: Occult papillary carcinoma of the thyroid. A "normal" finding in Finland. A systematic autopsy study. Cancer 56:531-538, 1985.
15. Folkman J, Kalluri R: Cancer without disease. Nature 427:787, 2004.
16. Hanahan D, Folkman J: Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 86:353-364, 1996.
17. Hasan J, Byers R, Jayson GC: Intratumoural microvessel density in human solid tumours. Br J Cancer 86:1566-1577, 2002.
18. Dvorak HF: Vascular permeability factor/ vascular endothelial growth factor: A critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. J Clin Oncol 20:4368-4380, 2002.
19. Patan S, Munn LL, Jain RK: Intussusceptive microvascular growth in a human colon adenocarcinoma xenograft: A novel mechanism of tumor angiogenesis. Microvasc Res 51:260-272, 1996.
20. Patan S, Munn LL, Tanda S, et al: Vascular morphogenesis and remodeling in a model of tissue repair: Blood vessel formation and growth in the ovarian pedicle after ovariectomy. Circ Res 89:723-731, 2001.
21. Patan S, Tanda S, Roberge S, et al: Vascular morphogenesis and remodeling in a human tumor xenograft: Blood vessel formation and growth after ovariectomy and tumor implantation. Circ Res 89:732-739, 2001.
22. Carmeliet P: Mechanisms of angiogenesis and arteriogenesis. Nat Med 6:389-395, 2000.
23. Jain RK, Schlenger K, Hockel M, et al: Quantitative angiogenesis assays: Progress and problems. Nat Med 3:1203-1208, 1997.
24. Jain RK: Molecular regulation of vessel maturation. Nat Med 9:685-693, 2003.
25. Isner JM: Myocardial gene therapy. Nature 115:234-239, 2002.
26. Rafii S, Lyden D, Benezra R, et al: Vascular and haematopoietic stem cells: Novel targets for anti-angiogenesis therapy? Nat Rev Cancer 2:826-835, 2002.
27. Jain RK, Duda DG: Role of bone marrow- derived cells in tumor angiogenesis and treatment. Cancer Cell 3:515-516, 2003.
28. Stoll BR, Migliorini C, Kadambi A, et al: A mathematical model of the contribution of endothelial progenitor cells to angiogenesis in tumors: Implications for antiangiogenic therapy. Blood 102:2555-2561, 2003.
29. Di Tomaso E, Capen D, Haskell A, et al: Mosaic tumor vessels: Cellular basis and ultrastructure of focal regions lacking endothelial cell markers. Cancer Res (in press).
30. Jain RK: Barriers to drug delivery in solid tumors. Sci Am 271:58-65, 1994.
31. Jain RK: Vascular and interstitial biology of tumors, in Abeleff M, Armitage J, Kastan M, et al (eds): Clinical Oncology, 3rd ed, chap 9, p 153. Philadelphia, Elsevier, 2004.
32. Bottaro DP, Liotta LA: Cancer: Out of air is not out of action. Nature 423:593-595, 2003.
33. Semenza GL: Targeting HIF-1 for cancer therapy. Nat Rev Cancer 3:721-732, 2003.
34. Jain RK, Safabakhsh N, Sckell A, et al: Endothelial cell death, angiogenesis, and microvascular function after castration in an androgen- dependent tumor: Role of vascular endothelial growth factor. Proc Natl Acad Sci USA 95:10820-10825, 1998.
35. Ferrara N, Gerber HP, LeCouter J: The biology of VEGF and its receptors. Nat Med 9:669-676, 2003.
36. Relf M, LeJeune S, Scott PA, et al: Expression of the angiogenic factors vascular endothelial cell growth factor, acidic and basic fibroblast growth factor, tumor growth factor beta-1, platelet-derived endothelial cell growth factor, placenta growth factor, and pleiotrophin in human primary breast cancer and its relation to angiogenesis. Cancer Res 57:963-969, 1997.
37. Sugimoto H, Hamano Y, Charytan D, et al: Neutralization of circulating vascular endothelial growth factor (VEGF) by anti-VEGF antibodies and soluble VEGF receptor 1 (sFlt- 1) induces proteinuria. J Biol Chem 278:12605- 12608, 2003.
38. Kasahara Y, Tuder RM, Taraseviciene- Stewart L, et al: Inhibition of VEGF receptors causes lung cell apoptosis and emphysema. J Clin Invest 106:1311-1319, 2000.
39. Baffert F, Thurston G, Rochon-Duck M, et al: Age-related changes in vascular endothelial growth factor dependency and angiopoietin-1-induced plasticity of adult blood vessels. Circ Res 94:984-992, 2004.
40. Zelzer E, Mamluk R, Ferrara N, et al: VEGFA is necessary for chondrocyte survival during bone development. Development 131:2161-2171, 2004.
41. Ratner M: Genentech discloses safety concerns over Avastin. Nat Biotechnol 22:1198, 2004.
42. Miller KD, Chap LI, Holmes FA, et al: Randomized phase III trial of capecitabine compared with bevacizumab plus capecitabine in patients with previously treated metastatic breast cancer. J Clin Oncol 23:792-799, 2005.
43. Ansiaux R, Baudelet C, Jordan BF, et al: Thalidomide radiosensitizes tumors through early changes in the tumor microenvironment. Clin Cancer Res 11:743-750, 2005.
44. Gazit Y, Baish JW, Safabakhsh N, et al: Fractal characteristics of tumor vascular architecture during tumor growth and regression. Microcirculation 4:395-402, 1997.
45. Hobbs SK, Monsky WL, Yuan F, et al: Regulation of transport pathways in tumor vessels: Role of tumor type and microenvironment. Proc Natl Acad Sci USA, 95:4607-4612, 1998.
46. Inai T, Mancuso M, Hashizume H, et al: Inhibition of vascular endothelial growth factor (VEGF) signaling in cancer causes loss of endothelial fenestrations, regression of tumor vessels, and appearance of basement membrane ghosts. Am J Pathol 165:35-52, 2004.
47. Kadambi A, Mouta Carreira C, Yun CO, et al: Vascular endothelial growth factor (VEGF)-C differentially affects tumor vascular function and leukocyte recruitment: Role of VEGF-receptor 2 and host VEGF-A. Cancer Res 61:2404-2408, 2001.
48. Lee CG, Heijn M, di Tomaso E, et al: Anti-vascular endothelial growth factor treatment augments tumor radiation response under normoxic or hypoxic conditions. Cancer Res 60:5565-5570, 2000.
49. Tong RT, Boucher Y, Kozin SV, et al: Vascular normalization by vascular endothelial growth factor receptor 2 blockade induces a pressure gradient across the vasculature and improves drug penetration in tumors. Cancer Res 64:3731-3736, 2004.
50. Wildiers H, Guetens G, De Boeck G, et al: Effect of antivascular endothelial growth factor treatment on the intratumoral uptake of CPT-11. Br J Can 88:1979-1986, 2003.
51. Winkler F, Kozin SV, Tong RT, et al: Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response to radiation: Role of oxygenation, angiopoietin-1, and matrix metalloproteinases. Cancer Cell 6:553-563, 2004.
52. Yuan F, Chen Y, Dellian M, et al: Timedependent vascular regression and permeability changes in established human tumor xenografts induced by an anti-vascular endothelial growth factor/vascular permeability factor antibody. Proc Natl Acad Sci USA 93:14765-14770, 1996.
53. Willett CG, Boucher Y, di Tomaso E, et al: Direct evidence that the VEGF-specific antibody bevacizumab has antivascular effects in human rectal cancer. Nat Med 10:145-147, 2004.
54. Morgan B, Thomas AL, Drevs J, et al: Dynamic contrast-enhanced magnetic resonance imaging as a biomarker for the pharmacological response of PTK787/ZK 222584, an inhibitor of the vascular endothelial growth factor receptor tyrosine kinases, in patients with advanced colorectal cancer and liver metastases: Results from two phase I studies. J Clin Oncol 21:3955-3964, 2003.
55. Soker S, Takashima S, Miao HQ, et al: Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 92:735- 745, 1998.
56. Bachelder RE, Crago A, Chung J, et al: Vascular endothelial growth factor is an autocrine survival factor for neuropilin-expressing breast carcinoma cells. Cancer Res 61:5736-40, 2001.
57. Li M, Yang H, Chai H, et al: Pancreatic carcinoma cells express neuropilins and vascular endothelial growth factor, but not vascular endothelial growth factor receptors. Cancer 101:2341-2350, 2004.
58. Izumi Y, Xu L, di Tomaso E, et al: Tumour biology: Herceptin acts as an antiangiogenic cocktail. Nature 416:279-280, 2002.
59. Donovan MJ, Lin MI, Wiegn P, et al: Brain derived neurotrophic factor is an endothelial cell survival factor required for intramyocardial vessel stabilization. Develop ment 127:4531-4540, 2000.
60. Duda DG, Jain RK: Pleiotropy of tissuespecific growth factors: From neurons to vessels via the bone marrow. J Clin Invest 115:596- 598, 2005.
61. Conrotto P, Valdembri D, Corso S, et al: Sema4D induces angiogenesis through Met recruitment by Plexin B1. Blood 2005. E-published ahead of print.
62. Torres-Vazquez J, Gitler AD, Fraser SD, et al: Semaphorin-plexin signaling guides patterning of the developing vasculature. Dev Cell 7:117-123, 2004.
63. Garkavtsev I, Kozin SV, Chernova O, et al: The candidate tumour suppressor protein ING4 regulates brain tumour growth and angiogenesis. Nature 428:328-332, 2004.
64. Wang B, Xiao Y, Ding BB, et al: Induction of tumor angiogenesis by Slit-Robo signaling and inhibition of cancer growth by blocking Robo activity. Cancer Cell 4:19-29, 2003.
65. Holden SN, Morrow M, O’Bryant C, et al: Correlative biological assays used to guide dose escalation in a phase I study of the antiangiogenic αVβ3 and αVβ5 integrin antagonist EMD 121974 (EMD) (abstract 110). Proc Am Soc Clin Oncol 21:28a, 2002.
66. Lu X, Le Noble F, Yuan L, et al: The netrin receptor UNC5B mediates guidance events controlling morphogenesis of the vascular system. Nature 432:179-186, 2004.












Late Hepatic Recurrence From Granulosa Cell Tumor: A Case Report
Granulosa cell tumors exhibit late recurrence and rare hepatic metastasis, emphasizing the need for lifelong surveillance in affected patients.


