Apoptosis Mechanisms: Implications for Cancer Drug Discovery
Defects in the regulation of apoptosis (programmed cell death) makeimportant contributions to the pathogenesis and progression of mostcancers and leukemias. Apoptosis defects also figure prominently inresistance to chemotherapy, radiotherapy, hormonal therapy, andimmune-based treatments. Apoptosis is caused by activation ofintracellular proteases, known as caspases, that are responsible directlyor indirectly for the morphologic and biochemical events thatcharacterize the apoptotic cell. Numerous proteins that regulate thesecell death proteases have been discovered, including proteins belongingto the Bcl-2, inhibitor of apoptosis, caspase-associated recruitmentdomain, death domain, and death effector domain families. Thesecaspase-regulating proteins provide mechanisms for linkingenvironmental stimuli to cell death responses or to maintenance of cellsurvival. Alterations in the expression and function of several apoptosisregulatinggenes have been demonstrated in cancer, suggesting targetsfor drug discovery. Knowledge of the molecular details of apoptosisregulation and the three-dimensional structures of apoptosis proteinshas revealed new strategies for identifying small-molecule drugs thatmay yield more effective treatments for malignancies. Apoptosisregulatinggenes are also beginning to find utility as targets for antisenseoligonucleotides.
ABSTRACT: Defects in the regulation of apoptosis (programmed cell death) makeimportant contributions to the pathogenesis and progression of mostcancers and leukemias. Apoptosis defects also figure prominently inresistance to chemotherapy, radiotherapy, hormonal therapy, andimmune-based treatments. Apoptosis is caused by activation ofintracellular proteases, known as caspases, that are responsible directlyor indirectly for the morphologic and biochemical events thatcharacterize the apoptotic cell. Numerous proteins that regulate thesecell death proteases have been discovered, including proteins belongingto the Bcl-2, inhibitor of apoptosis, caspase-associated recruitmentdomain, death domain, and death effector domain families. Thesecaspase-regulating proteins provide mechanisms for linkingenvironmental stimuli to cell death responses or to maintenance of cellsurvival. Alterations in the expression and function of several apoptosisregulatinggenes have been demonstrated in cancer, suggesting targetsfor drug discovery. Knowledge of the molecular details of apoptosisregulation and the three-dimensional structures of apoptosis proteinshas revealed new strategies for identifying small-molecule drugs thatmay yield more effective treatments for malignancies. Apoptosisregulatinggenes are also beginning to find utility as targets for antisenseoligonucleotides.Defects in the mechanisms ofprogrammed cell death(apoptosis) play importantroles in many aspects of tumor pathogenesisand progression. For example,apoptosis defects allow neoplasticcells to survive beyond their normallyintended life spans.[1] Thus, the needfor exogenous survival factors is subverted.Protection is provided againsthypoxia and oxidative stress as tumormass expands; and time is allowed foraccumulative genetic alterations thatderegulate cell proliferation, interferewith differentiation, promote angiogenesis,and increase cell motility andinvasiveness during tumor progression.In fact, apoptosis defects are recognizedas an important complementto proto-oncogene activation, becausemany deregulated oncoproteins thatdrive cell division also trigger apoptosis(eg, Myc, E1a, Cyclin-D1).[2] Similarly,defects in DNA repair and chromosomesegregation normally triggercell suicide as a defense mechanismfor eradicating genetically unstablecells; thus, apoptosis defects permitsurvival of the genetically unstablecells, providing opportunities for selectionof progressively more aggressiveclones.[3]Apoptosis defects also facilitatemetastasis by allowing epithelial cellsto survive in a suspended state, withoutattachment to an extracellular matrix.[4] These defects also promoteresistance to the immune system becausemany of the weapons that cytolyticT cells and natural killer cellsuse for attacking tumors depend on theintegrity of the apoptosis machinery.[5] Finally, cancer-associated defectsin apoptosis play a role inchemoresistance and radioresistance,increasing the threshold for cell deathand thereby requiring higher doses fortumor killing.[6] Thus, defective regulationof apoptosis is a fundamentalaspect of the biology of cancer.Because apoptosis defects permit awide variety of aberrant cellular behaviors,as exhibited in cancer cells,therapeutic strategies that negate theapoptosis advantage for tumors arepredicted to selectively kill cancercells as opposed to normal cells. Fundamentally,cancer cells should bemore dependent on apoptosis defensemechanisms than normal cells andthus should be proportionally moresensitive to interventions that targetapoptosis proteins and genes. To date,efforts to bring apoptosis-based strategiesinto animal models or humanclinical trials have provided supportfor this concept of selective vulnerabilityof neoplastic cells as opposed tonormal cells.


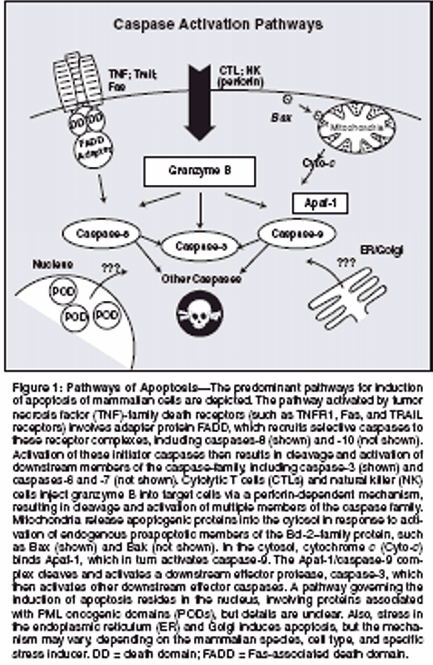
A solid knowledge base now existsabout the mechanisms of apoptosisregulation, the proteins involved, their3D structures, and biochemicalmechanisms. Over the past 2 decades,a clearer understanding has emergedof the defects in expression or functionof apoptosis-regulating genes andproteins relating to cancer. This informationcan now be exploited fordevising strategies for small-moleculedrug discovery toward the goal ofrevolutionary treatments for cancerand leukemia.Apoptosis PathwaysApoptosis is caused by proteasesknown as caspases, which stands forcysteine aspartyl-specific proteases.[7,8] Caspases constitute a familyof intracellular cysteine proteasesthat collaborate in proteolytic cascades,where caspases activate themselvesand each other.[9,10] Withinthese proteolytic cascades, caspasescan be positioned as either upstream"initiators" or downstream "effectors"of apoptosis.[11] Eleven caspases havebeen identified in the human genome.Several pathways for activatingcaspases probably exist, though detailsremain sketchy for some of them (Figure1). The simplest pathway is exploitedby cytolytic T cells and naturalkiller cells, which inject apoptosisinducingproteases, particularlygranzyme B, into target cells viaperforin channels.[12,13] Unlike thecaspases, granzyme B is a serine protease,but similar to the caspases,granzyme B specifically cleaves substratesat Asp residues. Granzyme Bis capable of cleaving and activatingmultiple caspases and some caspasesubstrates. Endogenous and viral inhibitorsof granzyme B have beenidentified, accounting for resistance tothis apoptotic inducer.[14-16]Another caspase-activation pathwayis represented by the tumor necrosisfactor (TNF)-family receptors.Of the approximately 30 known membersof the TNF family in humans, 8contain a so-called death domain intheir cytosolic tails.[17] Several ofthese death domain-containing TNFfamilyreceptors use caspase activationas a signaling mechanism, includingTNFR1/CD120a, Fas/APO1/CD95,DR3/Apo2/Weasle, DR4/TrailR1,DR5/TrailR2, and DR6. Ligation ofthese receptors at the cell surface resultsin the recruitment of several intracellularproteins, including certainprocaspases, to the cytosolic domainsof these receptors, forming a "deathinducingsignaling complex" (DISC)that triggers the activation of caspasesand leads to apoptosis.[18,19] Thespecific caspases summoned to theDISC are caspase-8 and, in somecases, caspase-10. These caspases containso-called death effector domainsin their N-terminal prodomains thatbind to a corresponding death effectordomain in the adapter protein, Fasassociateddeath domain (FADD), thuslinking them to the TNF-family deathreceptor complexes.Mitochondria also play importantroles in apoptosis, releasing cytochromec into the cytosol, which thencauses assembly of a multiproteincaspase-activating complex, referredto as the "apoptosome."[20,21] Thecentral component of the apoptosomeis Apaf-1, a caspase-activating proteinthat oligomerizes on binding cytochromec and that specifically bindsprocaspase-9. Apaf-1 and procaspase-9 interact via their caspase-associatedrecruitment domains (CARDs). Sucha CARD-CARD interaction plays importantroles in many steps in the pathwaysof apoptosis.The mitochondrial pathway forapoptosis is activated by myriadstimuli, including growth factor deprivation,oxidants, Ca2+ overload,DNA-damaging agents, and others.Mitochondria can also participate incell death pathways induced via TNFfamilydeath receptors, through crosstalkmechanisms involving proteinssuch as Bid, BAR, and Bap31.[22-25]However, mitochondrial (intrinsic)and death receptor (extrinsic) pathwaysfor the activation of caspases arefully capable of independent operationin most types of cells.[26] In additionto cytochrome c, mitochondria alsorelease several other proteins of relevanceto apoptosis, including endonucleaseG, AIF (an activator ofnuclear endonucleases), and inhibitorof apoptosis protein (IAP) antagonistsSmac (Diablo) and Omi (HtrA2).Pathways of apoptosis linked todamage in the endoplasmic reticulumand Golgi, as well as a pathway linkedto nuclear structures called PODs(PML oncogenic domains) or nuclearbodies, have also been described butare poorly characterized to date.


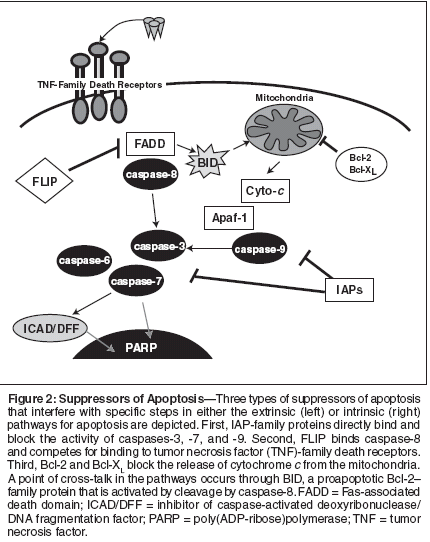
Suppressors of ApoptosisSeveral antagonists of the caspaseactivationpathways have been discovered,and multiple examples ofdysregulation of their expression orfunction in cancers have been obtained.Because our current knowledgeis greatest where the mitochondrial("intrinsic") and TNF-family deathreceptor ("extrinsic") pathways forapoptosis are concerned, most availableinformation about antagonistscenters on these two apoptotic pathways.In this article, three types ofapoptosis-suppressing proteins knownto be overexpressed in tumors, includingprostate cancers, are considered:IAPs, FLIP, and Bcl-2.Inhibitor of Apoptosis ProteinsInhibitor of apoptosis proteins representan evolutionarily conservedfamily of suppressors of apoptosis.Members of the IAP family, originallyidentified in baculoviruses, containone or more copies of a domain calledthe baculoviral IAP repeat (BIR).These BIR domains are sometimesaccompanied by other domains, includingRING domains, ubiquitinconjugatingenzyme folds (E2s), andNACHT-family nucleotide-bindingdomains. The human genome encodeseight IAP-family members: XIAP,cIAP1, cIAP2, Naip, Apollon (Bruce),ILP2 (Ts-IAP), ML-IAP (K-IAP;Livin), and Survivin.The BIR domains of several IAPfamilyproteins were originally shownby our laboratory to be responsible fordirectly binding and specifically inhibitingcaspases, thus identifyingIAPs as endogenous inhibitors of celldeath proteases.[27-31] Multipleother laboratories have confirmed andextended these findings, providingconclusive evidence that many IAPfamilyproteins operate as caspasesuppressors.[32-41] However, IAPsvary in the specific caspases they inhibit.For example, XIAP suppressesboth downstream effector caspasesthat operate at points of convergenceof apoptosis pathways and caspase-9, the apical protease in the mitochondrialpathway for apoptosis.[27,29,30]In contrast, ML-IAP is a potentsuppressor of only caspase-9. No examplesof IAP-mediated suppressionof proteases that operate in the upstreamportions of the apoptosis pathwayactivated by TNF-family receptorshave been found (Figure 2).Evidence of overexpression ofIAPs in cancer has been obtained, suggestinga role for these suppressors ofapoptosis in malignancy.[31,42] Forexample, the IAP-family memberSurvivin is overexpressed in most cancers[43] and has become a topic ofconsiderable attention for its dual roleas a regulator of cell division (chromosomesegregation and cytokinesis)and apoptosis.[44-46] Similarly, theIAP-family member ML-IAP is rarelyexpressed in normal tissues but isfound at elevated levels in melanomasand some renal cancers.[33,40,47]Moreover, XIAP has been reported byour group to be overexpressed in asubstantial proportion of acutemyelogenous leukemias, with higherlevels correlating with shorter remissiondurations and shorter overallpatient survival.[48] Evidence ofoverexpression of XIAP has also beenreported for renal and lung cancers[49,50]; overexpression of cIAP1has been associated with ovarian cancer.Chromosomal translocationsactivating cIAP2 are found in somelymphomas.[51] Thus, various IAPfamilyproteins are overexpressed inspecific types of cancer.However, more than one memberof the IAP family can be overexpressedsimultaneously by sometumors. For example, in prostatecancers, we found evidence thatprotein levels of XIAP, cIAP1, cIAP2,and Survivin can sometimes becomesimultaneously increased in tumors,[52] suggesting redundancy inexpression of these antiapoptotic proteins.We have also found evidence ofapparent simultaneous overexpressionof cIAP1, cIAP2, and Survivin in coloncancer (manuscript in preparation).The observation of overexpression ofmultiple IAP-family members impliesthat perhaps some aspects of theirregulation are shared.Indeed, during a screen of the NationalCancer Institute panel of 60human tumor cell lines, assessing IAPexpression at the messenger RNA(mRNA) and protein levels, we obtainedevidence that mRNA levels ofXIAP, cIAP1, and cIAP2 do not correlatewith their protein levels,[48]suggesting that posttranscriptionalregulation of these IAP-family proteinsis important. Interestingly, allthree of these IAP-family proteins containa RING domain that binds E2s(ubiquitin-conjugating enzymes), implyingthat alternations in the turnoverrate of IAP-family proteins may occurin cancers that overexpress multiplefamily members simultaneously.The functional importance ofoverexpressed IAPs for apoptosis suppressionin cancers has been supportedby antisense experiments.[53-57] Inthese experiments, knocking downexpression of Survivin, XIAP, or otherIAPs has been shown to induceapoptosis of tumor cell lines in cultureor to sensitize tumor cell lines toapoptosis induced by anticancerdrugs.[53-57] In contrast, gene knockoutstudies in mice imply that normalcells are possibly less dependent onIAPs than tumor cells because targeteddisruption of the genes for xiap, ciap1,and ciap2, both individually and incombination, produces little phenotype.[58; personal communication, T.Mak, 2004]Implications for Treatment
Taken together, these observationsimply that drugs that interfere with theaction of IAPs could be useful for thetreatment of cancer. Recently, a strategyfor devising small-molecule inhibitorsof IAPs has been suggestedby the discovery of natural antagonistsof IAPs.[35,38] Proteins such as Smacand Omi (HtrA2) have been shown tobind IAPs and suppress them, releasingcaspases to kill cells.[35,38,59] A7'mer peptide corresponding to the Nterminusof Smac is reported to besufficient to bind IAPs and block theirassociation with caspases.[60]Moreover, we have confirmed thatpeptides as short as tetramers can potentlyreverse caspase inhibition byIAPs, functioning in a stoichiometricmanner at micromolar concentrations.[61; unpublished data] By fusingmembrane-penetrating peptidesonto Smac or Omi peptides, it is possibleto induce apoptosis of cancer celllines in culture as well as to suppresstumor formation in xenograft modelsin mice.[62-65] Thus, these data provideproof-of-concept evidence thatsmall molecules that mimic the effectsof these IAP-binding peptides couldpotentially be exploited as drugs forcancer treatment.Drug Discovery Strategies
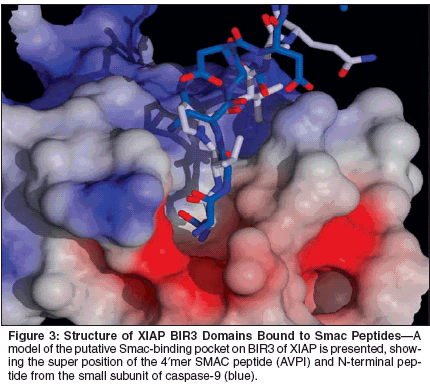
Structural analysis of the interactionsof IAPs with caspases and ofIAPs with Smac has helped to lay afoundation for such drug-discoveryefforts. First, our structure-functionstudies of IAP-family member XIAPshowed that, although this protein containsthree tandem BIR domains, asingle BIR is sufficient to bind andsuppress caspases. These studies demonstratedthat the BIR2 domain specificallyinhibits caspase-3 andcaspase-7, whereas the BIR3 domainof XIAP blocks the activity of caspase-9.[29,30] Thus, discrete domains inIAPs are responsible for binding andinhibiting caspases.Second, the 3D structure of theBIR3 domain complexed with Smacrevealed that the N-terminal 4 aminoacids of the mature Smac protein bindsin the same crevice normally occupiedby the N-terminus of the small subunitof caspase-9, thus suggestingcompletion for binding.[37,60,66,67]Consequently, small-molecule compoundsthat mimic the Smac 4?merpeptide should dislodge activecaspase-9 from BIR3, thus inducingapoptosis (Figure 3).The structural details regarding theinteraction of BIR2 of XIAP withcaspases and its relation to Smac areless clear due to poor atomic resolutionof the N-terminus of the smallsubunitof caspases-3 or -7 complexedwith BIR2, as determined by x-raycrystallography by scientists at ourinstitution and elsewhere.[32,39] Inthe crystal structure of the XIAPBIR2-caspase-3 complex, the NH2-terminus of the caspase-3 p10 subunitinteracts with the surface of BIR2,[32]which may be an artifact of crystallization.Though, to date, the mechanismof inhibition of XIAP by Smacremains unclear, modeling studiessuggest the presence of a similarSmac-binding pocket on BIR2.In addition to chemical inhibitorsof IAPs based on mimicking Smac,other strategies can also be envisionedand have begun to be exploited. Forexample, using an enzyme derepressionassay where screens were performedto identify compounds capableof dislodging XIAP fromcaspase-3 and restoring protease activity,we and other investigators haveidentified small-molecule antagonistsof XIAP.[68,69] These compoundstarget a non-Smac site on XIAP,which remains to be defined at thestructural level.Interestingly, in addition to Smacand Omi (HtrA2), other endogenousantagonists of IAPs have beenreported, including XAF1, NRAGE,and ARTS, which operate through analternative mechanism.[70-72] Thus,it is conceivable that the aforementionedsmall-molecule antagonistsof IAPs mimic one or more of theseendogenous antagonists of IAPs,a concept awaiting experimentaltesting.Fas-Associated Death Domain-Like Interleukin-1beta-Converting Enzyme InhibitingProtein (FLIP)The pathway of apoptosis triggeredby TNF-family death receptors is fundamentalto the mechanisms by whichcytolytic T cells attack and kill tumorcells.[73-78] Cytolytic T cells, naturalkiller cells, macrophages, and dendriticcells have been demonstrated toproduce one or more of the TNFfamilydeath ligands, such as FasL,TNF, or TRAIL. On binding their specificreceptors on susceptible targetcells, these receptors recruit procaspase-8 and/or -10 to the receptorcomplex, forming a DISC that resultsin the activation of caspases.[11,79]Perhaps not surprisingly, many tumorsdevelop resistance to this extrinsicpathway for apoptosis at some pointin their pathogenesis or progression,reducing or ablating their sensitivityto immune cell attack.[5]Multiple antagonists of the extrinsicpathway have been identified, includingseveral death effector domain-containing proteins that compete forbinding to the adapter proteins orprocaspases that participate in TNFfamilydeath receptor signaling, includingFLIP, BAR, and possiblyBap31.[80,81] Among them, FLIP hasreceived the most attention for its rolein producing Fas-resistant states intumor cells.[82,83]The FLIP protein is highly similarin its overall sequence to procaspases-8 and -10, containing tandem copiesof the death effector domain, followedby a pseudocaspase domain that lacksenzymatic activity. FLIP can promoteapoptosis in some circumstances.[84]However, for the most part, this proteinis antiapoptotic, forming complexeswith procaspase-8 and -10, andpreventing their effective activation, aswell as competing for binding toadapter proteins required for the recruitmentof caspases to receptors ofdeath-receptor complexes.[5,83]Overexpression of FLIP occurscommonly in cancers. Our laboratoryhas determined by antisense and genetransfer studies that FLIP is animportant determinant of resistance ofsome tumor cell lines to the inductionof apoptosis by TNF, Fas, andTRAIL.[85,86] Moreover, in a collaborativeeffort with other investigators,we have identified a class of compoundscalled synthetic triterpenoidsthat cause reductions in FLIP in multiplehuman tumor cell lines, correlatingwith the restoration of sensitivityto TRAIL-induced apoptosis.[85]Thus, small-molecule drugs that ablateexpression or function of FLIPrepresent an attractive approach to sensitizingtumor cells to TNF-familydeath ligands.The prototype triterpenoidshown to reduce the expression ofFLIP is CDDO (2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid). Atsubmicromolar levels, this compound,as well as selected analogs, reducesthe levels of FLIP protein by a mechanisminvolving ubiquitination andproteasome-dependent degradation ofthe FLIP protein without affectingFLIP mRNA levels.[85,87,88]CDDO has no effect on the levels ofmany other apoptosis proteins atsubmicromolar levels sufficient to reduceFLIP, including FADD, caspase-8, DAP3, XIAP, Bcl-2, Bcl-XL, Bax,Mcl-1, and Bak. However, the mechanismof this compound is presentlyundefined, and, therefore, it is likelythat other proteins besides FLIP areinvolved in its proapoptotic mechanism.Indeed, when employedat higher doses, CDDO and relatedcompounds are reported to have effectson a number of cancer-relevanttargets.[89]When tested on solid-tumor celllines at submicromolar concentrations,CDDO generally sensitizes only toTRAIL and Fas, but alone CDDO doesnot induce significant apoptosis. Appliedto primary leukemia cells, however,CDDO and related compoundsdemonstrate single-agent activity, inducingrobust apoptosis via a caspase-8-dependent mechanism, includingchemorefractory chronic lymphocyticleukemias and acute myelogenousleukemias.[87,90,91] Thus, by activatingthe extrinsic pathway, CDDO andrelated triterpenoids may providea pharmacologic route to bypassroadblocks to intrinsic pathway(mitochondrial) apoptosis, therebyachieving apoptotic destruction ofchemorefractory leukemias.




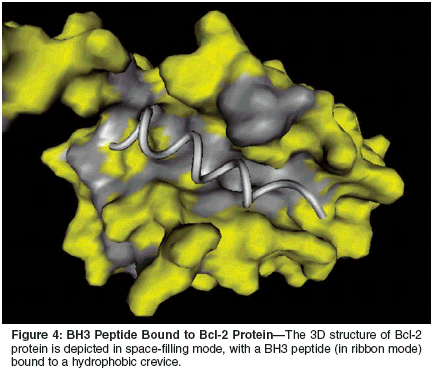
Biologic agents that trigger theextrinsic pathway are also being exploredfor their utility in cancer treatment.For example, an agonistic antibodythat activates TRAIL receptor-1(DR4) has been tested in phase I trialsof patients with malignancy. Recombinanttrimeric TRAIL protein has producedimpressive preclinical results inmouse models, either alone or in combinationwith chemotherapy, and alsomay soon enter clinical trials.[92]Bcl-2-Family ProteinsBcl-2 protein is the founding memberof a large family of apoptosisregulatingproteins that govern the intrinsicpathway of apoptosis.Overexpression of the antiapoptoticprotein Bcl-2 occurs in roughly halfof human cancers, contributing to resistanceto anticancer drugs, hormoneablative therapy, and radiotherapy.[1]Several homologs of Bcl-2 proteinhave been identified and characterized,with some functioning as blockers (n= 5 in humans) and others as promotersof cell death (n = 19 in humans),[93-95] comprising a gene familyof 25 members.[96; unpublisheddata].Alterations in the expressionof several members of the Bcl-2-family protein have been documentedin cancers, includingoverexpression of antiapoptotic membersand loss of expression of proapoptoticmembers.[97] Simultaneousoverexpression of more than one of thesix antiapoptotic members of the Bcl-2-family proteins can occur in somecancers, creating challenges with respectto overcoming roadblocks toapoptosis.Our prior analysis of prostate tumors,for example, revealed that levelsof antiapoptotic proteins Bcl-2,Bcl-XL, and Mcl-1 are commonly elevatedin advanced prostate cancers,whereas proapoptotic proteins Baxand Bak generally remain present athigh levels during progression of thesetumors to a hormone-independent,metastatic phenotype.[98] Similarfindings have been made for melanomas,which commonly overexpressproteins Bcl-2, Bcl-XL, and Mcl-1.[99]Bcl-2-family proteins operate asregulators of the mitochondriadependentpathway for apoptosis(intrinsic pathway). These proteinsgovern the permeability of the mitochondrialmembrane, dictatingwhether apoptogenic proteins such ascytochrome c are released into thecytosol.[21,95,100] One of the prominentmechanisms by which the mitochondrial(intrinsic) pathway forapoptosis cross-talks with the deathreceptor (extrinsic) pathway involvescaspase-8-mediated cleavage and activationof the proapoptotic Bcl-2 homologBid.Normally, Bid resides in the cytosolin a latent (inactive) state; however,on cleavage by caspase-8, this proteintranslocates to the outer membrane ofthe mitochondria, where it dimerizeswith other Bcl-2--family proteins, inducingthe release of cytochrome c andapoptosis.[95,101] Thus, in tumorcells where this Bid activation mechanismplays an important role in deathreceptor-mediated apoptosis (socalledtype II cells),[102,103]overexpression of either protein Bcl-2 or Bcl-XL has been shown to blockcell death induced by Fas, TNF, andTRAIL. Consequently, sensitivity oftumor cells to TNF-family cytokinescan potentially be improved by agentsthat reduce the expression or functionof Bcl-2/Bcl-XL.Many Bcl-2-family proteins physicallyinteract, forming homo- orheterodimers, whereby pro- andantiapoptotic members of this familyperform "hand-to-hand combat" inmaking cell life and death decisions.[104,105] The structural basisfor dimerization of Bcl-2-family proteinshas been elucidated using highfieldnuclear magnetic resonancemethods, revealing a hydrophobiccrevice on the surface of antiapoptoticproteins Bcl-2 and Bcl-XL that bindsan alpha-helical BH3 domain fromproapoptotic Bcl-2 family proteinssuch as Bax or Bak (Figure4).[106,107] Proof-of-concept experimentsperformed with BH3 peptideshave demonstrated suppression of proteinsBcl-2 or Bcl-XL, resulting in inductionof apoptosis or sensitizationof tumor cell lines to apoptosis in culture.[108-111]In addition, prototype nonpeptidylcompounds that compete for bindingto the BH3-binding pocket have beenidentified, enhancing sensitivity oftumor cell lines and cultured primaryleukemias to apoptotic stimuli (includinganticancer drugs), thus further validatingantiapoptotic Bcl-2-family proteinsBcl-2 and Bcl-XL as targets fordrug discovery.[112-117]Where tested, the compounds describedto date inhibit proteins Bcl-2,Bcl-XL, and (in some instances) otherantiapoptotic members of the Bcl-2family.[115,117] Thus, these compoundsmay afford advantagesover more targeted antisense-basedstrategies for overcoming apoptosis resistance,where redundancy is aproblem because of simultaneousoverexpression of more than oneantiapoptotic Bcl-2-family protein.However, because these compounds"hit" more targets, they may also proveto be more toxic to normal cells andtissues than antisense-based drugs.Thus, the therapeutic index of theseagents must be defined in vivo beforeclinical trials could be contemplated.Nevertheless, at least one smallmoleculeantagonist of protein Bcl-2has already been preliminarily evaluatedin human clinical trials, thoughits mechanism as an inhibitor of proteinBcl-2 was unknown at the timethe trials were initiated. This compound,gossypol, is a natural productidentified through Chinese herbalmedicine that interacts with the BH3-binding pocket of protein Bcl-2 withsubmicromolar affinity.[115] Gossypolundoubtedly has other targets besidesprotein Bcl-2; however, we andour collaborators have shown that gossypoldisplaces BH3 peptides fromantiapoptotic proteins Bcl-2, Bcl-XL,Bfl-1, and Bcl-B.[118] Semisyntheticanalogs of Gossypol are being evaluatedin preclinical studies to reduce thechemical reactivity of this compoundand thereby improve its safety andpharmacologic properties while retainingactivity against protein Bcl-2.[117]An alternative to small-moleculeantagonists that inhibit the Bcl-2 proteinis antisense DNA-based drugs thatsuppress the production of the Bcl-2protein.[119] These nuclease-resistant,synthetic, single-strand DNA moleculesbind cellular target mRNAs viaWatson-Crick base-pairing, leading toBcl-2 mRNA degradation by RNaseH-based mechanisms.[120] The Bcl-2 antisense drug oblimersen sodium(Genasense) hybridizes with the first18 nucleotides within the codingregion of Bcl-2 mRNAs, reducingthe expression of Bcl-2 proteinand thereby promoting apoptosis.Oblimersen sodium is in clinical testingfor several types of cancer.[121]ConclusionsKnowledge of the pathways ofapoptosis and of the mechanisms ofthe proteins that govern them is beginningto reveal a variety of targetsfor the discovery of cancer drugs. Detailedstructural analysis of apoptoticproteins and studies of their biochemicalmechanisms have suggested strategiesfor lead generation, resulting innumerous novel chemical entities withmechanism-based activity. Encouragingproof-of-principle data have beenprovided that help to validate severaltargets of apoptosis.Much work lies ahead, however, interms of optimizing the spectrum ofactivity of compounds that interactwith multiple members of apoptosisprotein families, improving the stabilityand pharmacologic properties ofthese compounds, establishing theiroptimal formulations for stability anddelivery, and elucidating attendantrate-limiting toxicities. Many of themost logical targets for promotingapoptosis of cancer and leukemia cellsare technically challenging and ofteninvolve either disrupting protein interactionsor altering gene expression, asopposed to traditional pharmaceuticalsthat typically target the active sites ofenzymes.Modern techniques of structurebaseddrug optimization render thistask feasible, but still challenging.Such targets require long-term commitments,often outstripping the usualdrug discovery and development cycleincorporated into the practices of pharmaceuticalcompanies. Long-termcommitments to research may createa new era in cancer therapy, where theintrinsic or acquired resistance of malignantcells to apoptosis can be pharmacologicallyreversed, reinstatingnatural pathways for cell suicide.There is good reason to suspect thatmalignant cells will be preferentiallysusceptible to restoration of apoptosissensitivity compared with normalcells. Cancer cells exhibit a wide varietyof abnormal behaviors andmolecular processes that normallywould trigger an apoptosis response,including cell-cycle checkpointdysregulation, oncogene activation,chromosome segregation defects, celldetachment from substratum, and outgrowthof blood supply (hypoxia).These defects render tumor cells moredependent on apoptosis-suppressinggenes and proteins, and thus withdrawingthis support from malignant cellsmay promote self-destruction of transformedcells while sparing normalcells.The full validity of this hypothesisawaits verification in human clinicaltrials. However, present insights fromanimal studies and current forays intothe clinic are encouraging. Apoptosisbasedstrategies for the discovery ofcancer drugs promise to yield effectivetherapies against cancer and meritfurther research support.
Disclosures:
Dr. Reed is a shareholderin Genta Incorporated.
References:
1.
Reed J: Dysregulation of apoptosis in cancer.J Clin Oncol 17:2941, 1999.
2.
Evan G, Littlewood T: A matter of lifeand cell death. Science 281:1317-1322, 1998.
3.
Ionov Y, Yamamoto H, Krajewski S, et al:Mutational inactivation of the pro-apoptoticgene BAX confers selective advantage duringtumor clonal evolution. Proc Natl Acad SciU S A 97:10872-10877, 2000.
4.
Frisch SM, Screaton RA: Anoikis mechanisms.Curr Opin Cell Biol 13:555-562, 2001.
5.
Tschopp J, Martinon F, Hofmann K:Apoptosis: Silencing the death receptors. CurrBiol 9:R381-R384, 1999.
6.
Makin G, Hickmann JA: Apoptosis andcancer chemotherapy. Cell Tissue Res 301:143,2000.
7.
Thornberry NA, Lazebnik Y: Caspases:Enemies within. Science 281:1312-1316, 1998.
8.
Cryns V, Yuan Y: Proteases to die for.Genes Dev 12:1551-1570, 1999.
9.
Thornberry NA, Rano TA, Peterson EP,et al: A combinatorial approach defines specificitiesof members of the caspase family andgranzyme B. J Biol Chem 272:17907-17911,1997.
10.
Karanewsky DS, Bai X, Linton SD, etal: Conformationally constrained inhibitors ofcaspase-1 (interleukin-1 beta converting enzyme)and of the human CED-3 homologuecaspase-3 (CPP32, apopain). Bioorg Med ChemLett 8:2757-2762, 1998.
11.
Salvesen GS, Dixit VM: Caspases: Intracellularsignaling by proteolysis. Cell91:443-446, 1997.
12.
Talanian R, Yang X, Turbov J, et al: Granule-mediated killing: Pathways for granzymeB initiated apoptosis. J Exp Med 186:1323-1331, 1997.
13.
Quan LT, Tewari M, O’Rourke K, et al:Proteolytic activation of the cell death proteaseYama/CPP32 by granzyme B. Proc Natl AcadSci U S A 93:1972-1976, 1996.
14.
Zhou Q, Krebs J, Snipas S, et al: Interactionof the baculovirus antiapoptotic proteinp35 with caspases: Specificity, kinetics, andcharacterization of the caspase/p35 complex.Biochemistry 37:10757-10765, 1998.
15.
Quan LT, Caputo A, Bleackley RC, etal: Granzyme B is inhibited by the cowpox virusserpin cytokine response modifier A. J BiolChem 270:10377-10379, 1995.
16.
Sun J, Ooms L, Bird CH, et al: A newfamily of 10 murine ovalbumin serpins includestwo homologs of proteinase inhibitor 8 and twohomologs of the granzyme B inhibitor (proteinaseinhibitor 9). J Biol Chem 272:15434-15441, 1997.
17.
Locksley RM, Killeen N, Lenardo MJ:The TNF and TNF receptor superfamilies: Integratingmammalian biology. Cell 104:487-501, 2001.
18.
Wallach D, Varfolomeev EE, MalininNL, et al: Tumor necrosis factor receptor andFas signaling mechanisms. Ann Rev Immunol17:331-367, 1999.
19.
Yuan J: Transducing signals of life anddeath. Curr Opin Cell Biol 9:247-251, 1997.
20.
Reed JC: Cytochrome c: Can’t live withit; Can’t live without it. Cell 91:559-562, 1997.
21.
Green DR, Reed JC: Mitochondria andapoptosis. Science 281:1309-1312, 1998.
22.
Gross A, Yin X-M, Wang K, et al:Caspase cleaved BID targets mitochondria andis required for cytochrome c release, whileBCL-XL prevents this release but not tumornecrosis factor-R1/Fas death. J Biol Chem274:1156-1163, 1999.
23.
Li H, Zhu H, Xu C, et al: Cleavage ofBID by caspase 8 mediates the mitochondrialdamage in the Fas pathway of apoptosis. Cell94:491-501, 1998.
24.
Zhang H, Xu Q, Krajewski S, et al: BAR:An apoptosis regulator at the intersection ofcaspase and bcl-2 family proteins. Proc NatlAcad Sci U S A 97:2597-2602, 2000.
25.
Ng FWH, Nguyen M, Kwan T, et al:p28 Bap31, a Bcl-2/Bcl-XL-and procaspase-8-associated protein in the endoplasmic reticulum.J Cell Biol 139:327-338, 1997.
26.
Vaux DL, Strasser A: The molecular biologyof apoptosis. Proc Natl Acad Sci U S A93:2239-2244, 1996.
27.
Deveraux QL, Takahashi R, SalvesenGS, et al: X-linked IAP is a direct inhibitor ofcell death proteases. Nature 388:300-304,1997.
28.
Roy N, Deveraux QL, Takashashi R, etal: The c-IAP-1 and c-IAP-2 proteins are directinhibitors of specific caspases. EMBO J16:6914-6925, 1997.
29.
Takahashi R, Deveraux QL, Tamm I, etal: A single BIR domain of XIAP sufficient forinhibiting caspases. J Biol Chem 273:7787-7790, 1998.
30.
Deveraux QL, Roy N, Stennicke HR, etal: IAPs block apoptotic events induced bycaspase-8 and cytochrome c by direct inhibitionof distinct caspases. EMBO J 17:2215-2223, 1998.
31.
Deveraux QL, Reed JC: IAP family proteins:Suppressors of apoptosis. Genes Dev13:239-252, 1999.
32.
Riedl SJ, Renatus M, SchwarzenbacherR, et al: Structural basis for the inhibition ofcaspase-3 by XIAP. Cell 104:791-800, 2001.
33.
Kasof GM, Gomes BC: Livin, a novelinhibitor-of-apoptosis (IAP) family member. JBiol Chem 276:3238-3246, 2001.
34.
Bratton SB, Walker G, Srinivasula SM,et al: Recruitment, activation and retention ofcaspases-9 and -3 by Apaf-1 apoptosome andassociated XIAP complexes. EMBO J 20:998-1009, 2001.
35.
Verhagen AM, Ekert PG, Pakusch M, etal: Identification of DIABLO, a mammalianprotein that promotes apoptosis by binding toand antagonizing IAP proteins. Cell 102:43-53, 2000.
36.
Sanna MG, de Silva Correia J, DucreyO, et al: IAP suppression of apoptosis involvesdistinct mechanisms: The TAK1/JNK1 signalingcascade and caspase inhibition. Mol CellBiol 22:1754-1766, 2002.
37.
Srinivasula SM, Hegde R, Saleh A, etal: A conserved XIAP-interaction motif incaspase-9 and Smac/DIABLO regulatescaspase activity and apoptosis. Nature 410:112-116, 2001.
38.
Du C, Fang M, Li Y, et al: SMAC, a mitochondrialprotein that promotes cytochromec-dependent caspase activation by eliminatingIAP inhibition. Cell 102:33-42, 2000.
39.
Huang Y, Park YC, Rich RL, et al: Structuralbasis of caspase inhibition by XIAP: Differentialroles of the linker versus the BIR domain.Cell 104:781-790, 2001.
40.
Vucic D, Stennicke HR, Pisabarro MT,et al: ML-IAP, a novel inhibitor of apoptosisthat is preferentially expressed in human melanomas.Curr Biol 10:1359-1366, 2000.
41.
LaCount DJ, Hanson SF, Schneider CL,et al: Caspase inhibitor P35 and inhibitor ofapoptosis Op-IAP block in vivo proteolyticactivation of an effector caspase at differentsteps. J Biol Chem 275:15657-15664, 2000.
42.
Prefontaine GG, Lemieux ME, Giffin W,et al: Recruitment of octamer transcription factorsto DNA by glucocorticoid receptor. MolCell Biol 18:3416-3430, 1998.
54.
Ambrosini G, Adida C, Altieri DC: Anovel anti-apoptosis gene, Survivin, expressedin cancer and lymphoma. Nat Med 3:917-921,1997.
44.
Altieri DC, Marchisio PC, Marchisio PC:Survivin apoptosis: An interloper between celldeath and cell proliferation in cancer. Lab Invest79:1327-1333, 1999.
45.
Reed J, Reed S: Survivin’ cell-separationanxiety. Nat Cell Biol 1:E199-E200, 1999.
46.
Reed JC, Bischoff JR: BIRinging chromosomesthrough cell division-and survivin’the experience. Cell 102:545-548, 2000.
47.
Lin JH, Deng G, Huang Q, et al: KIAP,a novel member of the inhibitor of apoptosisprotein family. Biochem Biophys Res Commun279:820-831, 2000.
48.
Tamm I, Kornblau SM, Segall H, et al:Expression and prognostic significance of IAPfamilygenes in human cancers and myeloidleukemias. Clin Cancer Res 6:1796-1803,2000.
49.
Hofmann HS, Simm A, Hammer A, etal: Expression of inhibitors of apoptosis (IAP)proteins in non-small cell human lung cancer.J Cancer Res Clin Oncol 128:554-560, 2002.
50.
Ferreira CG, van der Valk P, Span SW,et al: Assessment of IAP (inhibitor of apoptosis)proteins as predictors of response to chemotherapyin advanced non-small-cell lung cancerpatients. Ann Oncol 12:799-805, 2001.
51.
Motegi M, Yonezumi M, Suzuki H, etal: API2-MALT1 chimeric transcripts involvedin mucosa-associated lymphoid tissue typelymphoma predict heterogeneous products. AmJ Pathol 156:807-812, 2000.
52.
Krajewska M, Krajewski S, Turner B, etal: Elevated expression of inhibitor of apoptosisproteins (IAPs) in prostate cancer. Clin CancerRes 9:4914-4925, 2003.
53.
Chen J, Wu W, Tahir SK, et al: Downregulationof survivin by antisense oligonucleotidesincreases apoptosis, inhibits cytokinesisand anchorage-independent growth. Neoplasia2:235-241, 2000.
54.
Gordon GJ, Appasani K, Parcells JP, etal: Inhibitor of apoptosis protein-1 promotestumor cell survival in mesothelioma. Carcinogenesis23:1017-1024, 2002.
55.
Li F, Zhang D, Fujise K: Characterizationof fortilin, a novel antiapoptotic protein.J Biol Chem 276:47542-47549, 2001.
56.
Sasaki H, Sheng Y, Kotsuji F, et al:Down-regulation of X-linked inhibitor ofapoptosis protein induces apoptosis inchemoresistant human ovarian cancer cells.Cancer Res 60:5659-5666, 2000.
57.
Holcik M, Yeh C, Korneluk RG, et al:Translational upregulation of X-linked inhibitorof apoptosis (XIAP) increases resistance toradiation induced cell death. Oncogene19:4174-4177, 2000.
58.
Harlin H, Reffey SB, Duckett CS, et al:Characterization of XIAP-deficient mice. MolCell Biol 21:3604-3608, 2001.
59.
Suzuki Y, Imai Y, Nakayama H, et al: Aserine protease, HtrA2/Omi, is released fromthe mitochondria and interacts with XIAP, inducingcell death. Mol Cell 8:613-621, 2001.
60.
Chai J, Du C, Wu JW, et al: Structuraland biochemical basis of apoptotic activationby Smac/DIABLO. Nature 406:855-862, 2000.
61.
Huang Q, Deveraux QL, Maeda S, et al:Evolutionary conservation of apoptosis mechanisms:Lepidopteran and baculoviral inhibitorof apoptosis proteins are inhibitors of mammaliancaspase-9. Proc Natl Acad Sci U S A97:1427-1432, 2000.
62.
Deng Y, Lin Y, Wu X: TRAIL-inducedapoptosis requires Bax-dependent mitochondrialrelease of Smac/DIABLO. Genes Dev16:33-45, 2002.
63.
Guo F, Nimmanapalli R, ParanawithanaS, et al: Ectopic overexpression of second mitochondria-derived activator of caspases (Smac/DIABLO) or cotreatment with N-terminus ofSmac/DIABLO peptide potentiates epothiloneB derivative-(BMS 247550) and Apo-2L/TRAIL-induced apoptosis. Blood 99:3419-3426, 2002.
64.
Carson JP, Behnam M, Sutton JN, et al:Smac is required for cytochrome c-inducedapoptosis in prostate cancer LNCaP cells. CancerRes 62:18-23, 2002.
65.
Chauhan D, Hideshima T, Rosen S, etal: Apaf-1/cytochrome c-independent andSMAC-dependent induction of apoptosis inmultiple myeloma (MM) cells. J Biol Chem276:24453-24456, 2001.
66.
Liu Z, Sun C, Olejniczak ET, et al: Structuralbasis for binding of Smac/DIABLO to theXIAP BIR3 domain. Nature 408:1004-1008,2000.
67.
Wu G, Chai J, Suber TL, et al: Structuralbasis of IAP recognition by Smac/DIABLO. Nature 408:1008-1012, 2000.
68.
Schimmer AD, Welsh K, Pinilla C, et al:Small-molecule antagonists of apoptosis-suppressorXIAP exhibit broad anti-tumor activity.Cancer Cell 5:25-35, 2004.
69.
Wu TY, Wagner KW, Bursulaya B, et al:Development and characterization ofnonpeptidic small molecule inhibitors of theXIAP/caspase-3 interaction. Chem Biol 10:759-767, 2003.
70.
Liston P, Fong WG, Kelly NL, et al: Identificationof XAF1 as an antagonist of XIAPanti-caspase activity. Nat Cell Biol 3:128-133,2001.
71.
Jordan BW, Dinev D, LeMellay V, et al:NRAGE is an inducible IAP-interacting proteinthat augments cell death. J Biol Chem276:39985-39989, 2001.
72.
Gottfried Y, Rotm A, Lotan R, et al: Themitochondrial ARTS protein promotesapoptosis through targeting XIAP. EMBO J23:1627-1635, 2004.
73.
Lowin B, Mattman C, Hahne M, et al:Comparison of Fas(Apo-1/CD95)- andperforin-mediated cytotoxicity in primary Tlymphocytes. Int Immunol 8:57-63, 1996.
74.
Tschopp J, Hofmann K: Cytotoxic Tcells: More weapons for new targets? TrendsMicrobiol 4:91-94, 1996.
75.
Nagata S, Golstein P: The Fas death factor.Science 267:1449-1456, 1995.
76.
Braun MY, Lowin B, French I, et al:Cytotoxic T cells deficient in both functionalfas ligand and perforin show residual cytolyticactivity yet lose their capacity to induce lethalacute graft-versus-host disease. J Exp Med183:657-661, 1996.
77.
Hsieh MH, Korngold R: Differential useof FasL- and perforin-mediated cytolyticmechanisms by T-cell subsets involved in graftversus-myeloid leukemia responses. Blood96:1047-1055, 2000.
78.
Lewinsohn DM, Bement TT, Xu J, et al:Human purified protein derivative-specificCD4+ T cells use both CD95-dependent andCD95-independent cytolytic mechanisms.J Immunol 160:2374-2379, 1998.
79.
Schulze-Osthoff K, Ferrari D, Los M, etal: Apoptosis signaling by death receptors. EurJ Biochem 254:439-459, 1998.
80.
Krammer PH: CD95’s deadly missionin the immune system. Nature 407:789-795,2000.
81.
Griffith T, Chin W, Jackson G, et al: Intracellularregulation of TRAIL-inducedapoptosis in human melanoma cells. J Immunol161:2833-2840, 1998.
82.
Irmler M, Thome M, Hahne M, et al:Inhibition of death receptor signals by cellularFLIP. Nature 388:190-195, 1997.
83.
Tschopp J, Irmler M, Thome M: Inhibitionof Fas death signals by FLIPs. Curr OpinImmunol 10:552-558, 1998.
84.
Chang DW, Xing Z, Pan Y, et al: c-FLIPLis a dual function regulator for caspase-8 activationand CD95-mediated apoptosis. EMBOJ 21:3704-3714, 2002.
85.
Kim Y, Suh N, Sporn M, et al: An induciblepathway for degradation of FLIP proteinsensitizes tumor cells to TRAIL-inducedapoptosis. J Biol Chem 277:22320-22329,2002.
86.
Hyer ML, Sudarshan S, Kim Y, et al:Down-regulation of c-FLIP sensitizes DU145prostate cancer cells to fas-mediated apoptosis.Cancer Biol Ther 4:405-410, 2002.
87.
Suh W-S, Kim Y, Schimmer AD, et al:Synthetic triterpenoids activate a pathway forapoptosis in AML cells involving downregulationof FLIP and sensitization to TRAIL.Leukemia 17:2122-2129, 2003.
88.
Ikeda T, Nakata Y, Kimura F, et al: Inductionof redox imbalance and apoptosis inmultiple myeloma cells by the noveltriterpenoid 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid. Mol Cancer Ther 3:39-45,2004.
89.
Lapillonne H, Konopleva M, Tsao T, etal: Activation of peroxisome proliferatoractivatedreceptor gamma by a novel synthetictriterpenoid 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid induces growth arrest andapoptosis in breast cancer cells. Cancer Res63:5926-5939, 2003.
90.
Pedersen IM, Kitada S, Schimmer A, etal: The triterpenoid CDDO induces apoptosisin refractory CLL B-cells. Blood 100:2965-2972, 2002.
91.
Ito Y, Pandey P, Place A, et al: The noveltriterpenoid 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid induces apoptosis of humanmyeloid leukemia cells by a caspase-8-dependentmechanism. Cell Growth Differ 11:261-267, 2000.
92.
Ashkenazi A, Pai RC, Fong S, et al:Safety and antitumor activity of recombinantsoluble Apo2 ligand. J Clin Invest 104:155-162,1999.
93.
Reed JC: Bcl-2 family proteins.Oncogene 17:3225-3236, 1998.
94.
Adams J, Cory S: The Bcl-2 protein family:Arbiters of cell survival. Science 281:1322-1326, 1998.
95.
Gross A, McDonnell JM, Korsmeyer SJ:BCL-2 family members and the mitochondriain apoptosis. Genes Dev 13:1899-1911, 1999.
96.
Reed JC, Doctor K, Rojas A, et al: Comparativeanalysis of apoptosis and inflammationgenes of mice and humans. Genome Res13:1376-1388, 2003.
97.
Kitada S, Pedersen IM, Schimmer A, etal: Dysregulation of apoptosis genes in hematopoieticmalignancies. Oncogene 21:3459-3474,2002.
98.
Krajewska M, Krajewski S, Epstein JI,et al: Immunohistochemical analysis of bcl-2,bax, bcl-X and mcl-1 expression in prostatecancers. Am J Pathol 148:1567-1576, 1996.
99.
Selzer E, Schlagbauer-Wadl H, OkamotoI, et al: Expression of Bcl-2 family membersin human melanocytes, in melanoma metastasesand in melanoma cell lines. MelanomaRes 8:197-203, 1998.
100.
Kroemer G, Reed JC: Mitochondrialcontrol of cell death. Nat Med 6:513-519, 2000.
101.
Nagata S: Biddable death. Nat Cell Biol1:E143-E144, 1999.
102.
Scaffidi C, Fulda S, Srinivasan A, et al:Two CD95 (APO-1/Fas) signaling pathways.EMBO J 17:1675-1687, 1998.
103.
Yin X-M, Wang K, Gross A, et al: Biddeficientmice are resistant to Fas-inducedhepatocellular apoptosis. Nature 400:886-891,1999.
104.
Kelekar A, Thompson CB: Bcl-2-familyproteins-the role of the BH3 domain inapoptosis. Trends Cell Biol 8:324-330, 1998.
105.
Huang DC, Strasser A: BH3-only proteins-essential initiators of apoptotic cell death.Cell 103:839-842, 2000.
106.
Sattler M, Liang H, Nettesheim D, etal: Structure of Bcl-xL-Bak peptide complex:Recognition between regulators of apoptosis.Science 275:983-986, 1997.
107.
Petros AM, Medek A, Nettesheim DG,et al: Solution structure of the antiapoptoticprotein Bcl-2. Proc Natl Acad Sci U S A98:3012-3017, 2001.
108.
Simonen M, Keller H, Heim J: The BH3domain of Bax is sufficient for interaction ofBax with itself, with our family members andfor induction of apoptosis. Eur J Biochem249:85-91, 1997.
109.
Cosulich S, Worrall V, Hedge P, et al:Regulation of apoptosis by BH3 domains in acell-free system. Curr Biol 7:913-920, 1997.
110.
Holinger E, Chittenden T, Lutz R: BakBH3 peptides antagonize Bcl-xL function andinduce apoptosis through cytochrome c-independentactivation of caspases. J Biol Chem274:13298-13304, 1999.
111.
Wang J-L, Zhang Z-J, Choksi S, et al:Cell permeable Bcl-2 binding peptides: Achemical approach to apoptosis induction intumor cells. Cancer Res 60:1498-1502, 2000.
112.
Degterev A, Lugovskoy A, Cardone M,et al: Identification of small-molecule inhibitorsof interaction between the BH3 domainand Bcl-xL. Nat Cell Biol 3:173-182, 2001.
113.
Tung SP, Kim KM, Basanez G, et al:Antimycin A mimics a cell-death-inducingBcl-2 homology domain 3. Nat Cell Biol3:183-191, 2001.
114.
Wang J-L, Liu D, Zhang Z-J, et al:Structure-based discovery of an organic compoundthat binds Bcl-2 protein and inducesapoptosis of tumor cells. Proc Natl Acad SciU S A 97:7124-7129, 2000.
115.
Kitada S, Sareth S, Leone M, et al:Molecular basis for anticancer activity of Gossypol.J Med Chem 46:4259-4264, 2003.
116.
Leone M, Zhai D, Sareth S, et al: Cancerprevention by tea polyphenols is linked totheir direct inhibition of anti-apoptotic Bcl-2-family proteins. Cancer Res 63:8118-8121,2003.
117.
Becattini B, Kitada S, Leone M, et al:Rational design and real time in-cell detectionof the pro-apoptotic activity of a novelcompound targeting Bcl-Xl. Chem Biol11:389-395, 2004.
118.
Pellecchia M, Reed JC: Inhibition ofantiapoptotic Bcl-2 family proteins by naturalpolyphenols: New avenues for cancerchemoprevention and chemotherapy. CurrPharm Design 10:1387-1398, 2004.
119.
Reed JC, Stein C, Subasinghe C, et al:Antisense-mediated inhibition of BCL2 protooncogeneexpression and leukemic cell growthand survival: Comparisons of phosphodiesterand phosphorothioate oligodeoxynucleotides.Cancer Res 50:6565-6570, 1990.
120.
Klasa RJ, Gillum AM, Klem RE, et al:Oblimersen Bcl-2 antisense: Facilitatingapoptosis in anticancer treatment. AntisenseNucleic Acid Drug Dev 12:193-213, 2002.
121.
Buchele T: Proapoptotic therapy withoblimersen (bcl-2 antisense oligonucleotide)-Review of preclinical and clinical results[article in German].Onkologie 26(suppl7):60-69, 2003.












